
Table of Contents
Introduction: The Science Behind Cherries and Their Red Glow

A cherry’s red surface catches the eye like few other fruits can manage. That crimson shade is not just decoration. It represents a complex chemical strategy refined over millions of years. The pigments responsible for this coloration are anthocyanins, a class of flavonoid compounds that accumulate in the vacuoles of cherry cells. These molecules absorb certain wavelengths of light while reflecting others, creating the spectrum we perceive as red.
Anthocyanins respond to environmental cues in ways that reveal the intelligence of plant biochemistry. Their concentration shifts with light intensity, temperature fluctuations, and cellular pH levels. This responsiveness ties the cherry’s appearance directly to its survival needs. The red glow functions as more than visual appeal. It serves as a shield against radiation, a weapon against oxidative damage, and a signal to animals that might spread its seeds.
The six secrets explored here connect the cherry’s color to broader patterns in plant evolution and ecology. Each section examines a different aspect of how anthocyanins contribute to the cherry’s success as a species. The focus remains on biochemical mechanisms and ecological relationships, grounded in research rather than speculation.
Table 1: Comparison of Cherries with Other Rosaceae Fruits
| Fruit | Primary Pigments | Dispersal Strategy |
|---|---|---|
| Cherries | Anthocyanins (cyanidin-3-glucoside) | Bird dispersal via bright red coloration |
| Apples | Anthocyanins (cyanidin-3-galactoside) and carotenoids | Mixed dispersal by mammals and birds |
| Pears | Chlorophyll remnants, carotenoids, minimal anthocyanins | Mammalian dispersal with green-yellow signaling |
| Peaches | Carotenoids and anthocyanins in varying ratios | Mammalian dispersal with orange-red hues |
| Plums | High anthocyanins (peonidin, cyanidin derivatives) | Bird and mammal dispersal with purple-blue tones |
| Apricots | Carotenoids dominate, low anthocyanin content | Mammalian dispersal with orange coloration |
| Raspberries | Anthocyanins concentrated in drupelets | Bird dispersal via aggregate fruit structure |
| Strawberries | Anthocyanins (pelargonidin-3-glucoside) | Mixed dispersal with accessible achenes on surface |
1. Cherries and the Molecular Secret of Anthocyanins
Table 2: Key Anthocyanins in Cherry Varieties
| Anthocyanin Type | Chemical Structure Feature | Color Expression |
|---|---|---|
| Cyanidin-3-glucoside | Single glucose on 3-position | Deep red to purple-red |
| Cyanidin-3-rutinoside | Disaccharide (rutinose) attachment | Reddish-purple with stability |
| Peonidin-3-glucoside | Methoxylated cyanidin derivative | Purple-red with methoxy group |
| Pelargonidin-3-glucoside | Lacks B-ring hydroxyl group | Orange-red coloration |
| Cyanidin-3-sophoroside | Disaccharide (sophorose) attachment | Stable red pigment |
| Delphinidin derivatives | Additional B-ring hydroxylation | Blue-purple tones (rare in cherries) |
| Malvidin derivatives | Double methoxylation of delphinidin | Purple-blue (minimal in cherries) |
| Petunidin derivatives | Methoxylated delphinidin | Violet tones (trace amounts) |
Anthocyanins are part of a broader category of substances known as flavonoids. They form through a biosynthetic pathway that starts with phenylalanine and proceeds through several enzymatic steps. The most abundant anthocyanin in sweet cherries is cyanidin-3-glucoside, which accounts for roughly 80 to 90 percent of total anthocyanin content in many cultivars. This specific molecule creates the characteristic deep red that distinguishes ripe cherries from other fruits.
The chemical structure of anthocyanins determines their color expression. These molecules consist of an anthocyanidin backbone attached to sugar groups. The number and type of sugar attachments influence stability and hue. Cyanidin produces red to purple shades depending on cellular pH, while pelargonidin generates orange-red tones. Cherries concentrate primarily on cyanidin derivatives, which explains their consistent coloration across varieties.
Two genes play central roles in anthocyanin production within cherry tissues. The ANS gene encodes anthocyanidin synthase, an enzyme that converts colorless leucoanthocyanidins into colored anthocyanidins. The UFGT gene codes for UDP-glucose flavonoid 3-O-glucosyltransferase, which stabilizes anthocyanidins by attaching glucose molecules. Expression of these genes increases as cherries mature, triggered by developmental signals and environmental factors like temperature drops and increased light exposure.
Research on sweet cherry varieties has shown that anthocyanin accumulation begins during the color break stage of ripening. This process accelerates as fruits transition from yellow-green to red over a period of seven to ten days. Gene expression studies reveal that UFGT activity peaks during this window, correlating directly with visible pigment deposition. The timing ensures that cherries advertise their ripeness precisely when seeds have reached full maturity.
2. Cherries and the Shield Against Ultraviolet Radiation
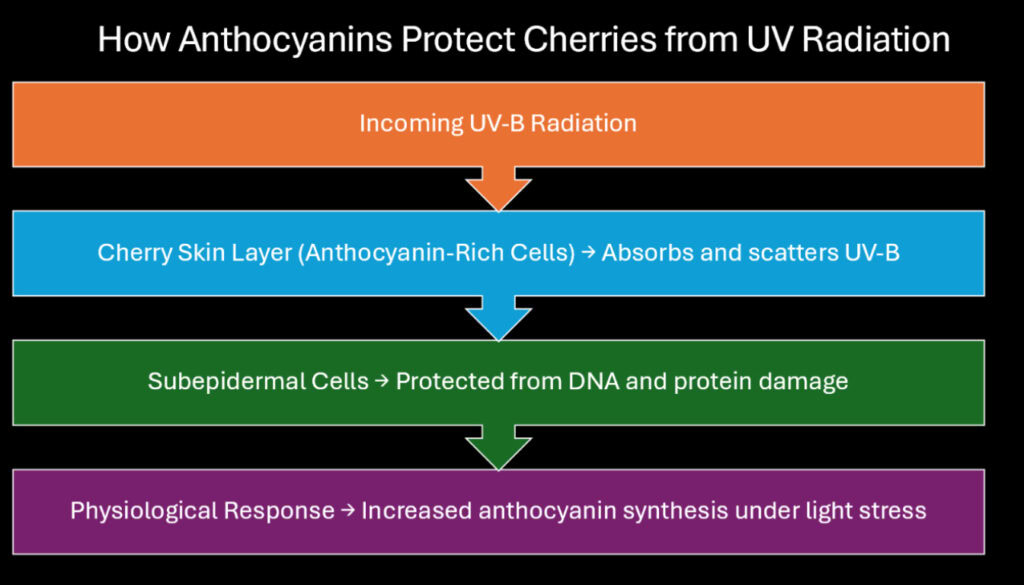
Ultraviolet radiation damages plant tissues at the molecular level. UV-B wavelengths between 280 and 315 nanometers penetrate cellular structures and cause mutations in DNA, disruption of proteins, and breakdown of membrane lipids. Cherries produce anthocyanins in their skin as a defensive response to this threat. These pigments absorb UV-B radiation before it reaches sensitive internal tissues, functioning as a natural sunscreen.
Studies on cherry cultivation at different elevations demonstrate this protective mechanism clearly. Trees grown above 1,000 meters show anthocyanin concentrations 30 to 40 percent higher than those at sea level. The increased UV exposure at altitude triggers enhanced pigment synthesis through light-responsive genetic pathways. This adaptation allows cherries to thrive in mountainous regions where radiation intensity would otherwise limit their range.
The absorption spectrum of anthocyanins matches UV-B wavelengths almost perfectly. Cyanidin-3-glucoside absorbs maximally around 280 nanometers, precisely where UV-B radiation poses the greatest danger. This specificity suggests evolutionary refinement over long periods. The pigment layer in cherry skin typically reaches depths of three to five cell layers, creating sufficient optical density to block approximately 95 percent of incoming UV-B radiation.
Research on cherry photobiology has measured how rapidly trees adjust pigment levels in response to changing light conditions. When shaded branches are suddenly exposed to full sun, anthocyanin synthesis increases within 48 hours. This rapid response indicates that cherries maintain constant surveillance of their light environment. The molecular sensors involved include photoreceptors called phytochromes and cryptochromes, which detect both UV and visible wavelengths.
Table 3: UV Protection Mechanisms in Cherry Tissues
| Protective Mechanism | Function |
|---|---|
| Anthocyanin absorption | Blocks 90-95% of UV-B radiation (280-315 nm) |
| Epidermal thickness | Creates physical barrier with 3-5 cell layers |
| Flavonoid accumulation | Provides additional UV screening compounds |
| Cuticle wax deposition | Reflects portion of UV radiation before absorption |
| DNA repair enzymes | Corrects UV-induced thymine dimers in cells |
| Antioxidant systems | Neutralizes free radicals generated by UV exposure |
| Heat shock proteins | Protects cellular machinery during radiation stress |
| Pigment density adjustment | Increases anthocyanins within 48 hours of UV exposure |
3. Cherries and Their Defense Against Oxidative Stress
Table 4: Oxidative Stress Response in Cherry Tissues
| Stress Factor | Cherry Response | Anthocyanin Role |
|---|---|---|
| High light intensity | Increases anthocyanin by 40-60% | Scavenges singlet oxygen from photosystems |
| Drought conditions | Elevates pigment synthesis in skin | Protects membranes from lipid peroxidation |
| Temperature extremes | Accelerates anthocyanin accumulation | Stabilizes proteins against denaturation |
| Pathogen attack | Induces phenolic and anthocyanin production | Neutralizes pathogen-generated ROS |
| Mechanical damage | Triggers localized pigment synthesis | Prevents oxidative damage at wound sites |
| Ripening metabolism | Coordinates with climacteric respiration | Maintains cellular integrity during senescence |
| Nutrient deficiency | Modifies anthocyanin ratios | Compensates for reduced enzyme antioxidants |
| Ozone exposure | Increases flavonoid production | Detoxifies ozone-derived free radicals |
Oxidative stress occurs when reactive oxygen species accumulate faster than cells can neutralize them. These molecules include superoxide radicals, hydrogen peroxide, and hydroxyl radicals. They form naturally during photosynthesis and respiration, but environmental stressors like drought, heat, and high light intensity accelerate their production. Left unchecked, reactive oxygen species destroy lipid membranes, denature proteins, and fragment nucleic acids.
Anthocyanins function as direct scavengers of these dangerous molecules. Their chemical structure contains multiple hydroxyl groups that donate electrons to reactive oxygen species, converting them into stable, harmless compounds. Studies measuring antioxidant capacity in cherry extracts have found that anthocyanin-rich fractions neutralize free radicals with efficiency comparable to synthetic antioxidants like butylated hydroxytoluene.
The location of anthocyanins within cells enhances their protective effectiveness. These pigments accumulate in vacuoles, which occupy up to 90 percent of mature cell volume. This positioning places them throughout the cytoplasm’s periphery, where they intercept reactive oxygen species before those molecules can damage vital organelles like mitochondria and chloroplasts. The concentration gradient ensures that oxidative molecules encounter anthocyanins almost immediately upon formation.
Research on cherry ripening has documented how anthocyanin synthesis correlates with periods of maximum oxidative stress. During the color break stage, when respiration rates peak and cells undergo rapid metabolic changes, anthocyanin concentrations increase by factors of five to ten. This timing suggests that pigment production serves a protective function during vulnerable developmental phases. Field studies comparing stressed and unstressed cherry trees confirm that drought or heat exposure triggers accelerated anthocyanin accumulation.
4. Cherries and the Battle Against Herbivores
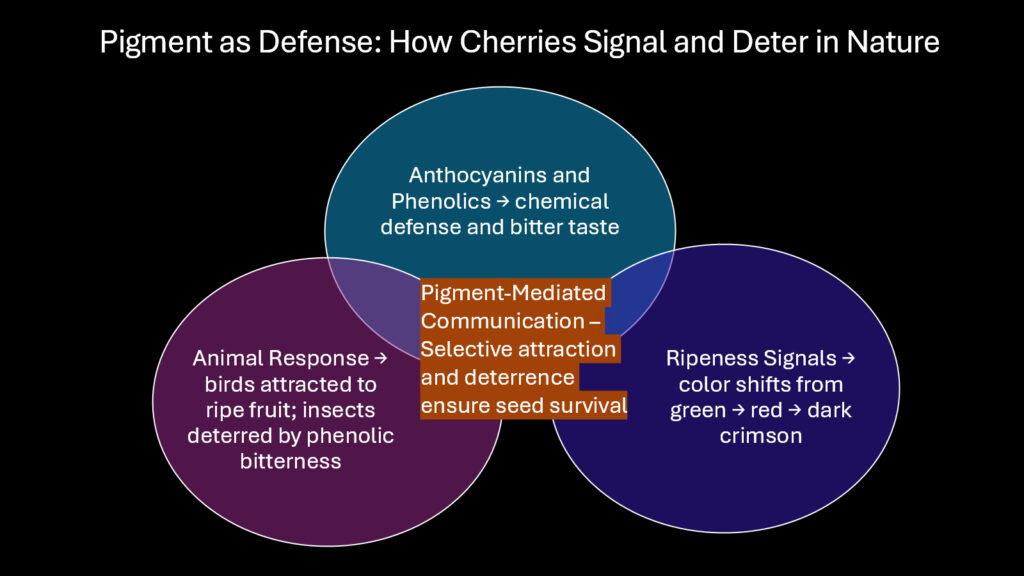
The relationship between cherries and herbivores involves complex chemical signaling. While ripe cherries attract seed dispersers through color and sweetness, unripe fruits deploy defensive compounds that discourage premature consumption. Anthocyanins contribute to this defense indirectly through their association with other phenolic compounds. The biosynthetic pathways that produce anthocyanins also generate tannins, flavonols, and hydroxycinnamic acids, many of which taste bitter or act as digestive inhibitors.
Studies on cherry palatability have measured how anthocyanin concentration correlates with other defensive metabolites. Unripe cherries contain high levels of chlorogenic acid and condensed tannins alongside low anthocyanin levels. As fruits mature and anthocyanins increase, these defensive compounds decrease proportionally. This shift transforms the fruit from unpalatable to attractive precisely when seeds have developed the capacity for germination.
The red coloration itself may serve as an honest signal of chemical defense in certain contexts. Research on fruit color evolution suggests that some herbivores learn to associate deep red or purple hues with high concentrations of secondary metabolites. For cherries, this creates a window of protection during early development when fruits are most vulnerable. The color change during ripening then signals a transition from defended to available, guiding herbivore behavior in ways that benefit the plant.
Insect herbivores respond differently to cherry pigmentation than mammalian herbivores do. Many insects lack red-sensitive photoreceptors and perceive ripe cherries as dark or black against green foliage. This visual limitation may reduce insect damage to ripe fruits while birds, which see red vividly, continue to locate and consume them readily. The differential visibility creates a selective filter that favors seed dispersers over seed predators.
Table 5: Chemical Defense Compounds in Cherry Development
| Development Stage | Primary Defense Strategy |
|---|---|
| Green stage (pre-veraison) | High tannins, low sugars, chlorogenic acid dominance |
| Color break initiation | Transition from tannins to anthocyanins begins |
| Early red stage | Moderate anthocyanins, decreasing astringency |
| Full red ripeness | Maximum anthocyanins, minimal defensive compounds |
| Post-harvest senescence | Oxidation of phenolics, loss of protective function |
| Unripe fruit taste | Bitter and astringent due to condensed tannins |
| Ripe fruit taste | Sweet with minimal bitterness, high palatability |
| Seed maturity correlation | Chemical shift matches seed viability timeline |
5. Cherries and the Language of Color for Pollinators and Dispersers
Table 6: Visual Ecology of Cherry-Disperser Interactions
| Disperser Type | Color Perception | Cherry Visibility |
|---|---|---|
| Songbirds (thrushes, waxwings) | Tetrachromatic with UV sensitivity | Excellent red detection against foliage |
| Mammals (rodents, deer) | Dichromatic vision | Moderate visibility, less contrast |
| Bees and wasps | UV-blue-green trichromatic | Poor red detection, see as dark |
| Butterflies and moths | Broad spectrum including red | Variable detection depending on species |
| Ants | UV-blue dichromatic | Minimal red perception |
| Human observers | Trichromatic RGB vision | Strong red-green contrast |
| Birds of prey | Enhanced tetrachromatic | Superior color discrimination |
| Fruit bats (if present) | Limited color vision | Rely on scent more than color |
Color perception varies dramatically across animal taxa. Birds possess four types of cone cells in their retinas, allowing them to see into the ultraviolet range and distinguish color variations invisible to mammals. Most mammals have only two cone types, giving them dichromatic vision similar to human red-green colorblindness. Insects typically see ultraviolet, blue, and green wavelengths but lack red sensitivity. Cherries have evolved coloration that speaks primarily to their most effective seed dispersers: birds.
The reflectance spectrum of ripe cherries shows peak reflectance around 650 to 680 nanometers, corresponding to deep red wavelengths. This matches the sensitivity range of avian red-sensitive cones almost perfectly. Studies using spectrometry have confirmed that cherries stand out prominently against green foliage when viewed through bird-vision models. The contrast between red fruit and green leaves creates a visual target that birds can detect from considerable distances.
Research on seed dispersal effectiveness has demonstrated that birds preferentially select red fruits over green, yellow, or orange alternatives when all are equally nutritious. This preference appears hardwired through evolutionary history. Bird-dispersed plant lineages show a strong bias toward red and black fruits, while mammal-dispersed species favor yellows, oranges, and browns. Cherries fit firmly within the bird-dispersed category, both in color and in seed structure.
The timing of color change in cherries coordinates with bird migration and breeding patterns in many regions. Sweet cherries ripen in late spring and early summer, when resident bird populations are feeding nestlings and require high-energy food sources. The availability of ripe cherries during this critical period strengthens the mutualistic relationship between plant and disperser. Birds consume fruits for their sugars and lipids while inadvertently transporting seeds to new locations through their digestive systems.
6. Cherries and the Influence of Climate on Their Red Glow
Table 7: Environmental Factors Affecting Cherry Pigmentation
| Environmental Factor | Optimal Range | Effect on Anthocyanins |
|---|---|---|
| Daytime temperature | 22-28°C | Supports biosynthesis without heat stress |
| Nighttime temperature | 10-15°C | Enhances enzyme activity and pigment stability |
| Light intensity | Full sun (>800 μmol/m²/s) | Maximizes photosynthate availability for pigments |
| UV-B radiation | Natural levels increase with altitude | Triggers defensive anthocyanin accumulation |
| Soil pH | 6.0-6.5 (slightly acidic) | Maintains optimal cellular pH for color expression |
| Water availability | Moderate deficit stress | Concentrates pigments without damaging tissues |
| Altitude | 500-1500 meters | Combines cool temps, high UV, and light intensity |
| Growing season length | 90-120 days for maturation | Allows complete anthocyanin development |
Temperature influences anthocyanin synthesis through multiple mechanisms. Cool nights during the ripening period promote pigment accumulation by enhancing enzyme activity in the biosynthetic pathway. Research on sweet cherry cultivation has found that diurnal temperature fluctuations of 10 to 15 degrees Celsius optimize color development. Constant warm temperatures inhibit anthocyanin production even when light exposure remains adequate.
Light intensity determines the overall capacity for anthocyanin synthesis. Cherries growing in full sun develop deeper coloration than those in partial shade, even when both receive identical temperature regimes. Photosynthesis provides the carbon skeletons and energy needed for flavonoid biosynthesis, creating a direct link between light availability and pigment production. Studies measuring fruit color as a function of canopy position show that sun-exposed fruits contain 40 to 70 percent more anthocyanins than shaded fruits.
Soil pH affects anthocyanin stability and expression through its influence on cellular pH. Anthocyanins are pH-sensitive molecules that shift color based on the acidity of their environment. In acidic conditions below pH 3, they appear bright red. As pH rises toward neutral, they fade to purple and eventually colorless. Cherry varieties grown in alkaline soils sometimes develop paler coloration due to this pH sensitivity, though genetic factors remain the primary determinant of final color.
Altitude combines several climatic factors that intensify anthocyanin production. Higher elevations expose cherries to greater UV radiation, lower temperatures, and increased diurnal temperature variation. All three factors stimulate pigment synthesis through different pathways. Comparative studies between lowland and highland cherry orchards demonstrate that elevation differences of 500 meters can result in anthocyanin concentration increases of 20 to 35 percent, producing noticeably darker fruits.
Conclusion: The Enduring Brilliance of Cherries in Nature’s Design
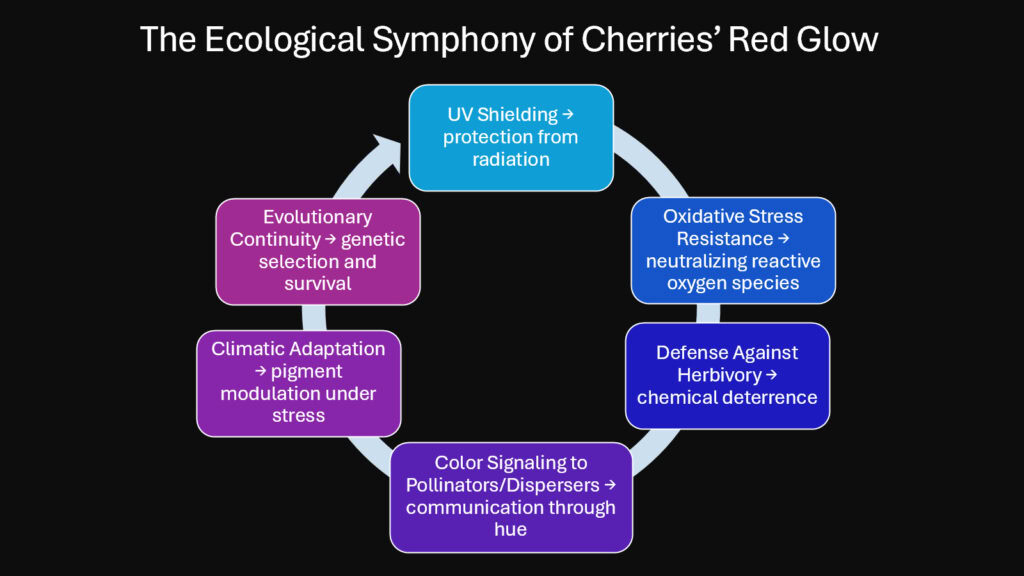
The red glow of cherries emerges from a convergence of molecular precision and ecological necessity. Anthocyanins function simultaneously as pigments, protectors, and communicators within a single fruit. Each role reinforces the others through biochemical pathways refined across millions of years. The UV shield protects tissues while signaling ripeness. The antioxidant defense maintains cellular integrity during the metabolic demands of maturation. The color language attracts precisely those animals capable of carrying seeds to favorable germination sites.
This integration reveals how evolution solves multiple problems with a single innovation. The genes that produce anthocyanins respond to light, temperature, and stress in ways that optimize fruit quality across varying environments. A cherry grown at high altitude develops deeper coloration than one at sea level, but both achieve the same fundamental goals: protecting developing seeds and ensuring their dispersal once mature. The system adapts without losing its core function.
The study of cherry pigmentation illuminates broader principles in plant biology. Color in nature rarely serves only aesthetic purposes. It encodes information about chemistry, ecology, and evolutionary history. When we examine cherries’ red surface through the lens of biochemistry, we see not just beauty but a sophisticated survival strategy written in molecular language. That glow represents millions of years of environmental conversation between plants and their world, a dialogue continuing in every orchard and wild cherry tree today.
Table 8: Summary of Anthocyanin Functions in Cherry Biology
| Function Category | Primary Mechanism | Ecological Outcome |
|---|---|---|
| UV protection | Absorption of 280-315 nm radiation | Prevents DNA damage in fruit tissues |
| Oxidative defense | Free radical scavenging via hydroxyl groups | Maintains cellular integrity during ripening |
| Visual signaling | Red wavelength reflection (650-680 nm) | Attracts bird dispersers effectively |
| Herbivore deterrence | Association with bitter tannins in unripe fruit | Reduces premature seed predation |
| Thermal regulation | Heat absorption and dissipation | Moderates temperature extremes in tissues |
| Ripeness indication | Concentration increase during maturation | Coordinates dispersal with seed viability |
| Drought response | Enhanced production under water stress | Protects tissues from desiccation damage |
| Altitude adaptation | Increased synthesis at high elevations | Compensates for enhanced UV exposure |


